Clock Tutorial #7: Circadian Organization in Mammals
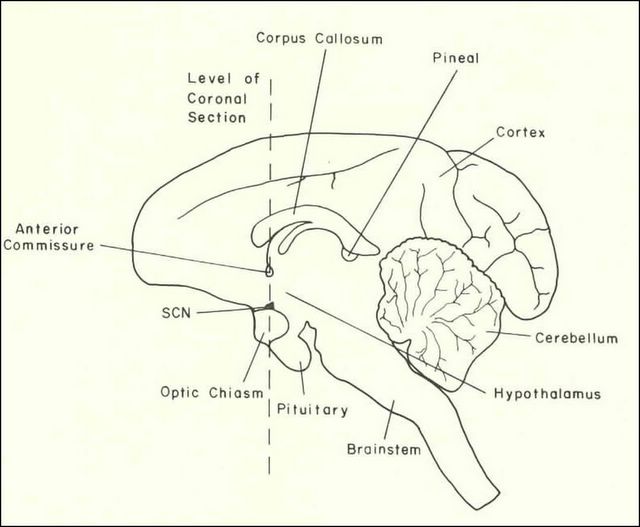
The principal mammalian circadian pacemaker is located in the suprachiasmatic nuclei (SCN) of the hypothalamus. The general area was first discovered in 1948 by Curt Richter who systematically lesioned a number of endocrine glands and brain areas in rats. The only time he saw an effect on circadian rhythms was when he lesioned a frontal part of hypothalamus (which is at the base of the brain) immediatelly above the optic chiasm (the spot where two optic nerves cross). Later studies in the 1970s narrowed the area to a small pair of nuclei, each composed of about 10,000 neurons. Similar studies (lesions) were performed in a number of other species of mammals, confirming that SCN is the pacemaker in all mammals.

[Image: Rat Brain]
A serendipitous discovery of a circadian period mutant (Tau-mutant) by Martin Ralph, then in Mike Menaker's lab, provided an opportunity to further test the hypothesis that SCN is, indeed, the pacemaker, and not just a way-station for the circadian information (as was still a possible alternative explanation of the lesion studies). The wild-type Golden hamsters have an endogenous period of almost precisely 24 hours. The homozygous tau-mutants exhibit a period of about 20 hours (while heterozygotes have an intermediate period of about 22 hours). Ralph transplantated SCNs between wild-type and Tau-mutant hamsters. What he saw was that the period of the overt rhythms of the hosts was always equal to the period of the transplanted SCN of the donor, i.e., a mutant hamster with a wild-type graft exhibited a 24-hour rhythm, while a wild-type mutant with implanted mutant SCN showed a periodicity of 20 hours. He rightly concluded that the SCN contained within itself all the information needed for the proper functioning of the circadian system, including the most basic property of the rhythm - its period - thus the SCN is, by definition, the pacemaker of the hamster circadian system.

Wild-Type Hamster

Tau-Mutant Hamster

Results of Transplantations
Each SCN is, itself, a multi-oscillator system as individual SCN neurons can show different endogenous periodicities in vitro. In vivo, the individual neurons are coupled together to produce a single circadian output. Recent studies indicate that some of the coupling is not synaptic (via neurotransmitters) but electrical (via "gap junctions"). More recent studies uncovered an internal division within each nucleus in what are termed "core" and "shell", each with slightly different properties, one with slightly shorter innate period than the other, and one recieving the majority of the neural input from the eye.

The eye is the only route for the information about external light conditions into the mammalian circadian system. The photic (light) input to the SCN comes from the ganglion cells of the retina via a direct mono-synaptic retinohypothalamic tract (i.e., the ganglion cell of the retina sends one long process all the way to the SCN, so the signal is NOT transduced via a chain of several successive neural cells). Non-visual photoreception, involved in circadian entrainment, pupillary reflex and control of mood, appears to be mediated differently from vision. Although photoreceptor cells (rods and cones) appear to play a role, most attention recently has been given to a small subset of retinal ganglion cells that project directly to the SCN and have been shown to be directly photo-sensitive. The identity of the photopigment used by these cells remains a subject of much debate, with melanopsin and cryptochrome being the major candidates. As persuasive data have been reported supporting an important role for each of these pigments, it is likely that both are involved, playing specific yet somewhat overlapping roles in non-visual photoreception.

[Image - Mammalian Retina]
The output of the clock is coupled to various effector areas of the brain including other areas of the hypothalamus, as well as to the superior cervical ganglia which provide adrenergic innervation of the pineal gland. Driven by the daily rhythm of adrenergic stimulation, the pineal synthesizes and secretes its hormone (melatonin) into the bloodstream during the night, but not during the day. In mammals, the profile of melatonin secretion is used for the interpretation of daylength in the photoperiodic response. There is no endogenous rhythmicity in the mammalian pineal - all pineal rhythms are driven by neural inputs from the SCN. The retina, on the other hand, is a site of a circadian clock but the retinal clock appears to drive rhythms within the eye itself and it does not influence the rest of the circadian system.
[Image - Primate brain]
Apparently, all cells in mammalian body contain circadian clocks. These cells, in contrast to cells of the SCN, are called peripheral clocks or oscillators. Peripheral circadian oscillators cannot maintain rhythmicity in the absence of the SCN. When placed in a dish, peripheral clocks may undergo a few oscillations before becoming arrhythmic. In contrast, SCN cells cycle indefinitely in vitro. In addition, peripheral oscillators are not directly light sensitive, so the only way they can be entrained is via neural or hormonal signal driven by the pacemaker in the SCN. A number of humoral signals have been hypothesized to play this role, including cortisol and melatonin. The dynamics of entrainment of peripheral clocks appears to vary between the tissues, e.g., liver cells being much slower to reset to a new light-dark schedule than cells in some other organs. This observation can potentially explain jet-lag as a result of internal desynchronization between various peripheral oscillators.
Sex differences have been noted in mammalian circadian systems and a number of studies have shown an interaction between the circadian and the reproductive systems. The role of circadian rhythms in the measurement of daylength as an environmental signal for seasonal reproduction has been demonstrated quite decisively in mammals. An involvement of the circadian system in the timing of ovulation (primarily in rodents) is not as clear, although some persuasive data have been published. Effects of implants of steroid hormones have been seen in female rodents, both castrated and intact, with slight increases in period effected by estradiol. This effect is blocked by simultaneous application of progesterone and is also seen in embryonically feminized males.
Archives/Categories: Clock Tutorials
Tags: circadian
[Image: Rat Brain]
A serendipitous discovery of a circadian period mutant (Tau-mutant) by Martin Ralph, then in Mike Menaker's lab, provided an opportunity to further test the hypothesis that SCN is, indeed, the pacemaker, and not just a way-station for the circadian information (as was still a possible alternative explanation of the lesion studies). The wild-type Golden hamsters have an endogenous period of almost precisely 24 hours. The homozygous tau-mutants exhibit a period of about 20 hours (while heterozygotes have an intermediate period of about 22 hours). Ralph transplantated SCNs between wild-type and Tau-mutant hamsters. What he saw was that the period of the overt rhythms of the hosts was always equal to the period of the transplanted SCN of the donor, i.e., a mutant hamster with a wild-type graft exhibited a 24-hour rhythm, while a wild-type mutant with implanted mutant SCN showed a periodicity of 20 hours. He rightly concluded that the SCN contained within itself all the information needed for the proper functioning of the circadian system, including the most basic property of the rhythm - its period - thus the SCN is, by definition, the pacemaker of the hamster circadian system.
Wild-Type Hamster
Tau-Mutant Hamster
Results of Transplantations
Each SCN is, itself, a multi-oscillator system as individual SCN neurons can show different endogenous periodicities in vitro. In vivo, the individual neurons are coupled together to produce a single circadian output. Recent studies indicate that some of the coupling is not synaptic (via neurotransmitters) but electrical (via "gap junctions"). More recent studies uncovered an internal division within each nucleus in what are termed "core" and "shell", each with slightly different properties, one with slightly shorter innate period than the other, and one recieving the majority of the neural input from the eye.
The eye is the only route for the information about external light conditions into the mammalian circadian system. The photic (light) input to the SCN comes from the ganglion cells of the retina via a direct mono-synaptic retinohypothalamic tract (i.e., the ganglion cell of the retina sends one long process all the way to the SCN, so the signal is NOT transduced via a chain of several successive neural cells). Non-visual photoreception, involved in circadian entrainment, pupillary reflex and control of mood, appears to be mediated differently from vision. Although photoreceptor cells (rods and cones) appear to play a role, most attention recently has been given to a small subset of retinal ganglion cells that project directly to the SCN and have been shown to be directly photo-sensitive. The identity of the photopigment used by these cells remains a subject of much debate, with melanopsin and cryptochrome being the major candidates. As persuasive data have been reported supporting an important role for each of these pigments, it is likely that both are involved, playing specific yet somewhat overlapping roles in non-visual photoreception.
[Image - Mammalian Retina]
The output of the clock is coupled to various effector areas of the brain including other areas of the hypothalamus, as well as to the superior cervical ganglia which provide adrenergic innervation of the pineal gland. Driven by the daily rhythm of adrenergic stimulation, the pineal synthesizes and secretes its hormone (melatonin) into the bloodstream during the night, but not during the day. In mammals, the profile of melatonin secretion is used for the interpretation of daylength in the photoperiodic response. There is no endogenous rhythmicity in the mammalian pineal - all pineal rhythms are driven by neural inputs from the SCN. The retina, on the other hand, is a site of a circadian clock but the retinal clock appears to drive rhythms within the eye itself and it does not influence the rest of the circadian system.
[Image - Primate brain]
Apparently, all cells in mammalian body contain circadian clocks. These cells, in contrast to cells of the SCN, are called peripheral clocks or oscillators. Peripheral circadian oscillators cannot maintain rhythmicity in the absence of the SCN. When placed in a dish, peripheral clocks may undergo a few oscillations before becoming arrhythmic. In contrast, SCN cells cycle indefinitely in vitro. In addition, peripheral oscillators are not directly light sensitive, so the only way they can be entrained is via neural or hormonal signal driven by the pacemaker in the SCN. A number of humoral signals have been hypothesized to play this role, including cortisol and melatonin. The dynamics of entrainment of peripheral clocks appears to vary between the tissues, e.g., liver cells being much slower to reset to a new light-dark schedule than cells in some other organs. This observation can potentially explain jet-lag as a result of internal desynchronization between various peripheral oscillators.
Sex differences have been noted in mammalian circadian systems and a number of studies have shown an interaction between the circadian and the reproductive systems. The role of circadian rhythms in the measurement of daylength as an environmental signal for seasonal reproduction has been demonstrated quite decisively in mammals. An involvement of the circadian system in the timing of ovulation (primarily in rodents) is not as clear, although some persuasive data have been published. Effects of implants of steroid hormones have been seen in female rodents, both castrated and intact, with slight increases in period effected by estradiol. This effect is blocked by simultaneous application of progesterone and is also seen in embryonically feminized males.
Archives/Categories: Clock Tutorials
Tags: circadian
posted by Bora Zivkovic at 8:13 PM
![]()
![]()






0 Comments:
Post a Comment
<< Home