Photoperiodism - Models and Experimental Approaches
In the previous ClockTutorials post, I wrote about the adaptive function of seasonality. Here, I will focus on photoperiodic time measurement - mostly on conceptual models and experimental designs invented to test those models. In the next installment, I will concentrate on actual physiology of photoperiodism, particularly in mammals, and then apply that to the human affliction - the Seasonal Affective Disorder (SAD, or 'Winter Blues').
Timely prediction of seasonal periods of weather conditions, food availability or predator activity is crucial for survival of many species. Although not the only parameter, the changing length of the photoperiod ('daylength') is the most predictive environmental cue for the seasonal timing of physiology and behavior, most notably for timing of migration, hibernation and reproduction. While rising spring temperatures may vary from year to year, the gradual increase in daylength is uniform and precise. Still, some cold-blooded organisms also respond to thermoperiod (oscillations between increasing duration of warmth during the days and decreasing duration of cold temperature during the nights) and some modulate their photoperiodic response by overall temperature levels.
Models of Photoperiodism
Three main models for photoperiodic time measurement have been proposed.
Garner and Allard, in 1920, discovered photoperiodism in plants. Soon afterwards, they and other researchers confirmed that changes in daylength, in absence of any other clues, trigger flowering in a number of plant species. Some respond to lengthening days in the spring, others to shortening days of the fall. Similar findings in insects, mammals and birds soon followed. The way these early researchers conceptualized photoperiodic response was later named the "hourglass model".
The hourglass model assumes the gradual accumulation of a chemical product in the organism. A certain quantity of this chemical is necessary to trigger a physiological response (e.g., flowering in plants, or growth of gonads in mammals). The threshold is reached if the product is not first degraded. It may be degraded by dark and only accumulates during the light phase or it may accumulate during dark and be degraded by light. If the light (or the dark) is long enough threshold is reached and a physiological response, such as maturation of the reproductive system, is initiated. This model argues against the involvement of the circadian clock in the photoperiodic response because the hourglass lacks endogenous rhythmicity and must be reset or "turned over" by the light cycle each day:
 The involvement of the circadian system was, however, detected in almost all species studied to date, so the hourglass mechanism is unlikely to have evolved in real organisms. However, its historical primacy, its simplicity which appeals to our sense of parsimony, and its usefulness as a hypothesis to test against, make the hourglass model an integral part of the scientific repertoire in the study of photoperiodism.
The involvement of the circadian system was, however, detected in almost all species studied to date, so the hourglass mechanism is unlikely to have evolved in real organisms. However, its historical primacy, its simplicity which appeals to our sense of parsimony, and its usefulness as a hypothesis to test against, make the hourglass model an integral part of the scientific repertoire in the study of photoperiodism.
The external coincidence model was proposed by one of the pioneers of chronobiology, Erwin Bunning, in 1936. This model proposed the existence of a circadian rhythm of photoperiodic photosensitivity (CRPP) in which most of the night-phase is sensitive to light, while the day-phase is photoinsensitive. As the day gets longer in spring, light starts illuminating the photosensitive phase and triggers the physiological or behavioral response:
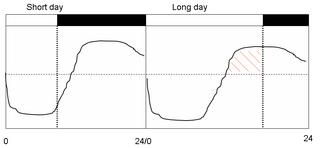 In this model light has a dual effect: it entrains the rhythm of photosensitivity and also acts as the stimulus (that is stimulates a photoperiodic response) if the light falls on a photosensitive phase of the rhythm of photoperiodic sensitivity. This model is termed an "external coincidence" model because it requires the coincidence (hatched red) of an external stimulus (light) with an internal rhythm of sensitivity to light.
In this model light has a dual effect: it entrains the rhythm of photosensitivity and also acts as the stimulus (that is stimulates a photoperiodic response) if the light falls on a photosensitive phase of the rhythm of photoperiodic sensitivity. This model is termed an "external coincidence" model because it requires the coincidence (hatched red) of an external stimulus (light) with an internal rhythm of sensitivity to light.
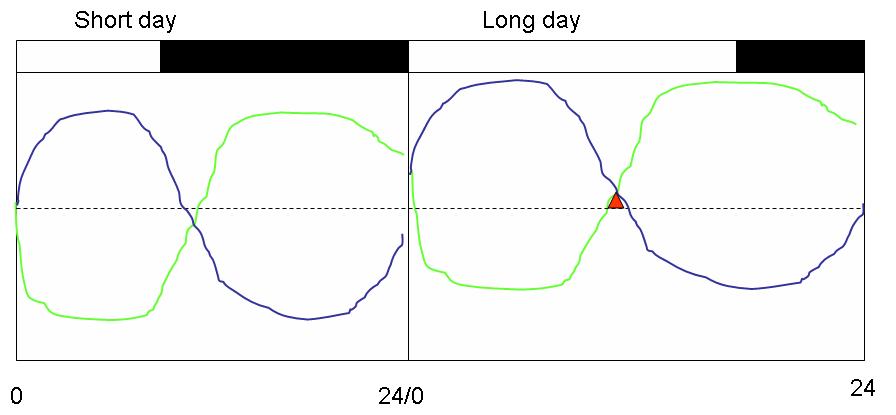
This dual role of light is not necessary if one considers the "internal coincidence" model, proposed by Colin Pittendrigh and Dorothea Minis in 1964. In this model, the light's only role is to entrain the circadian system. At the time this model was proposed, it was becoming apparent that multicellular organisms house more than one circadian pacemaker. Each of the oscillators will behave differently under the influence of the light-dark cycles, and assume different phase-relationships with the entraining cycle.
Pittendrigh and Minis proposed that changing photoperiods may alter the internal phase-relationships between two or more rhythms, bringing them into permissive (red) or inhibitory modes. For example, secretion of a hormone has to coincide with the availability of its receptors at the target tissue, or with the absence of the enzyme that metabolizes it. In this model, therefore, light must control only the internal phase-relationships between multiple circadian rhythms:
 Experimental Evidence
Experimental Evidence
Several experimental protocols have been developed to test circadian involvement in the photoperiodic response. Positive results of these experiments eliminate the hourglass model, thus confirm the involvement of the circadian system in photoperiodic time measurement. It was always hoped that, in the process, these experiments would also be able to distinguish between the internal and external coincidence models.
These protocols involve the use of night-break light pulses (i.e., skeleton photoperiods, see the "Entrainment" series of posts), resonance light cycles, and non-24 hour cycles ("T-cycle experiment") of light and dark.
Skeleton photoperiods mimic the full photoperiods with the light pulses at the times of dawn and dusk only. Although the total time per day that the animal spends in light is quite small, the photoperiodic response is elicited if the night-break pulse (or the second of the two skeleton pulses) falls during the photoinductive phase of the circadian cycle.
However, in skeleton photoperiods, it is often impossible to mimic long photoperiods as the circadian system tends to entrain to the shorter of the two possible interpretations of the skeleton photoperiods. Also, there is often an additional complication of phase-shifting effects of both pulses.
Resonance cycles are light-dark regimes in which the period T (L+D) is a multiple of 12 hours (e.g., LD 6:18, LD 6:30, LD 6:42, LD 6:54, etc.). If the circadian system is involved, every other light pulse will fall onto the photosensitive phase and induce a photoperiodic response in all cycles which are odd multiples of 12 hours (e.g., LD6:18, LD 6:42), while in cycles which are even multiples of 12 hours (e.g., LD 6:30, LD 6:54) the photoperiodic response will not be seen because every light pulse falls only during the insensitive phase of the rhythm of photoperiodic photosensitivity. In resonance cycles, the long periods of darkness may theoretically counter the stimulatory effects of the rare light pulses.
In T-cycle experiments, the period (T) of the LD cycle is different from 24 hours and varies within the limits of entrainment. During entrainment, the phase of the pacemaker is shifted or reset each cycle by an amount (Df) equal to the difference between the freerunning period (t) and the period (T) of the entraining cycle ( t - T = Df). The phase angle (y) between the onset of light and the onset of circadian activity will be characteristic of each T-cycle and can be calculated from the phase-response curve (PRC).
The PRC describes the phase-shifting behavior (lengths of advances or delays) of a circadian system elicited by a specific environmental perturbation (e.g., a pulse of light of defined length, intensity and wavelength) as a function of circadian timing of that perturbation. As a general rule, light pulses given around early subjective night elicit phase-delays, while those given late in the subjective night result in phase-advances.
In LD cycles of different period lengths, the phase angles between the onset of circadian activity and the onset of the light phase of the cycle will be different and predictable from the PRC. Accordingly, on some T-cycles the circadian system will be entrained in such a way that the light pulse illuminates a portion of the photoinducible phase (fi) and result in a measurable physiological photoperiodic response. If the internal coincidence model is correct, the entraining LD cycle will control the phases of different circadian outputs differently, thus changing the phase relationships between two or more circadian rhythms within the organism. Light-dark cycles of certain period lengths (T) will then bring the relevant rhythms into a phase relationship that results in the stimulation of the photoperiodically controlled physiological system.
The T-cycle protocol does not have the same difficulties in interpretation as the other two protocols. Only one light pulse per cycle is used and it has both an entraining and a potential photostimulatory effect. Utilizing different period-length cycles spanning the limits of entrainment of the organism, the light pulse can be brought into every possible phase-relationship with the circadian system. As the animals assume stable entrainment, the light pulse and the circadian system will assume a different phase-angle in each T-cycle. Presumably, in some of the cycles the light will repeatedly illuminate the photoperiodically photosensitive phase leading to the physiological response, while in other T-cycles, the light will be coincident only with the subjective day, leading to the lack of stimulation.
If an internal coincidence mechanism is operating, it is likely that the T-cycles will place the two circadian oscillators in all possible phase-relationships, leading to the stimulation in some and inhibition of response in other values of T. In theory, the two models may have different predictions as to which Ts will be inductive and which not.
So, what's the evidence?
In many organisms short days inhibit while long days stimulate various seasonal responses of physiology and behavior. Various experimental paradigms, including skeleton photoperiods and resonance cycles, demonstrated that it is not the total amount of light, but the precise temporal placing of the light that induces the photoperiodic response, giving the circadian system a role in the measurement and interpretation of daylength.
In a large number of species, including both invertebrates and vertebrates, as well as plants, these protocols have proven the involvement of the circadian system in photoperiodic time measurement, e.g., resonance cycles in male rosefinches, skeleton photoperiods in the rain quail Coturnix coromendelica, T-cycles and resonance cycles in golden hamsters and house finches, etc.
On the other hand, there are some organisms in which the results of all these protocols are not so conclusive. For instance, T-cycle and night-break studies in blackheaded buntings indicate the dependence of the reproductive response on the period (T) of the cycle and on the length of the light pulse within the cycle. In starlings, effects of light intensity, wavelength and pulse length can be seen even in 24 hour cycles.
Although the involvement of the circadian system has been documented in many species, it has not yet been possible to discriminate between the external and the internal coincidence mechanism for photoperiodic time measurement. So far, results of skeleton photoperiod, resonance cycle and T-cycle experiments could be interpreted equally well with both models.
Moreover, in some species it was even difficult to differentiate between the hourglass and circadian models. The vetch aphid (Megoura viciae) is probably the most notorious example. A mathematical model, though, was developed that succesfully describes photoperiodic responses in all of the hundreds of arthropod species that were studied over the past century. The fact that this mathematical model is consistent with all the experimental data on the vetch aphid suggests that it may also use its circadian clock to measure photoperiod, it's just that its clock's putput is of a very small amplitude.
Another species that was difficult to figure out was American chameleon (Anolis carolinensis). Initial studies suggested that hourglass model was operating. However, later work by Linda Hyde utilizing T-cycles showed that the circadian clock is indeed involved in photoperiodic time measurement in this species.
Finally, in Japanese quail (Coturnix japonica), results of skeleton photoperiods were positive, resonance cycles suggest some involvement of the clock but not the complete response, and the T-cycles were negative. This is also a species with a low-amplitude circadian system (i.e., it has a Type 0 PRC), so it is likely that clock is involved but the final demonstration is still waiting for the next creative experimenter to provide.
Category: Clock Tutorials
Timely prediction of seasonal periods of weather conditions, food availability or predator activity is crucial for survival of many species. Although not the only parameter, the changing length of the photoperiod ('daylength') is the most predictive environmental cue for the seasonal timing of physiology and behavior, most notably for timing of migration, hibernation and reproduction. While rising spring temperatures may vary from year to year, the gradual increase in daylength is uniform and precise. Still, some cold-blooded organisms also respond to thermoperiod (oscillations between increasing duration of warmth during the days and decreasing duration of cold temperature during the nights) and some modulate their photoperiodic response by overall temperature levels.
Models of Photoperiodism
Three main models for photoperiodic time measurement have been proposed.
Garner and Allard, in 1920, discovered photoperiodism in plants. Soon afterwards, they and other researchers confirmed that changes in daylength, in absence of any other clues, trigger flowering in a number of plant species. Some respond to lengthening days in the spring, others to shortening days of the fall. Similar findings in insects, mammals and birds soon followed. The way these early researchers conceptualized photoperiodic response was later named the "hourglass model".
The hourglass model assumes the gradual accumulation of a chemical product in the organism. A certain quantity of this chemical is necessary to trigger a physiological response (e.g., flowering in plants, or growth of gonads in mammals). The threshold is reached if the product is not first degraded. It may be degraded by dark and only accumulates during the light phase or it may accumulate during dark and be degraded by light. If the light (or the dark) is long enough threshold is reached and a physiological response, such as maturation of the reproductive system, is initiated. This model argues against the involvement of the circadian clock in the photoperiodic response because the hourglass lacks endogenous rhythmicity and must be reset or "turned over" by the light cycle each day:
 The involvement of the circadian system was, however, detected in almost all species studied to date, so the hourglass mechanism is unlikely to have evolved in real organisms. However, its historical primacy, its simplicity which appeals to our sense of parsimony, and its usefulness as a hypothesis to test against, make the hourglass model an integral part of the scientific repertoire in the study of photoperiodism.
The involvement of the circadian system was, however, detected in almost all species studied to date, so the hourglass mechanism is unlikely to have evolved in real organisms. However, its historical primacy, its simplicity which appeals to our sense of parsimony, and its usefulness as a hypothesis to test against, make the hourglass model an integral part of the scientific repertoire in the study of photoperiodism.The external coincidence model was proposed by one of the pioneers of chronobiology, Erwin Bunning, in 1936. This model proposed the existence of a circadian rhythm of photoperiodic photosensitivity (CRPP) in which most of the night-phase is sensitive to light, while the day-phase is photoinsensitive. As the day gets longer in spring, light starts illuminating the photosensitive phase and triggers the physiological or behavioral response:
 In this model light has a dual effect: it entrains the rhythm of photosensitivity and also acts as the stimulus (that is stimulates a photoperiodic response) if the light falls on a photosensitive phase of the rhythm of photoperiodic sensitivity. This model is termed an "external coincidence" model because it requires the coincidence (hatched red) of an external stimulus (light) with an internal rhythm of sensitivity to light.
In this model light has a dual effect: it entrains the rhythm of photosensitivity and also acts as the stimulus (that is stimulates a photoperiodic response) if the light falls on a photosensitive phase of the rhythm of photoperiodic sensitivity. This model is termed an "external coincidence" model because it requires the coincidence (hatched red) of an external stimulus (light) with an internal rhythm of sensitivity to light.This dual role of light is not necessary if one considers the "internal coincidence" model, proposed by Colin Pittendrigh and Dorothea Minis in 1964. In this model, the light's only role is to entrain the circadian system. At the time this model was proposed, it was becoming apparent that multicellular organisms house more than one circadian pacemaker. Each of the oscillators will behave differently under the influence of the light-dark cycles, and assume different phase-relationships with the entraining cycle.
Pittendrigh and Minis proposed that changing photoperiods may alter the internal phase-relationships between two or more rhythms, bringing them into permissive (red) or inhibitory modes. For example, secretion of a hormone has to coincide with the availability of its receptors at the target tissue, or with the absence of the enzyme that metabolizes it. In this model, therefore, light must control only the internal phase-relationships between multiple circadian rhythms:
 Experimental Evidence
Experimental EvidenceSeveral experimental protocols have been developed to test circadian involvement in the photoperiodic response. Positive results of these experiments eliminate the hourglass model, thus confirm the involvement of the circadian system in photoperiodic time measurement. It was always hoped that, in the process, these experiments would also be able to distinguish between the internal and external coincidence models.
These protocols involve the use of night-break light pulses (i.e., skeleton photoperiods, see the "Entrainment" series of posts), resonance light cycles, and non-24 hour cycles ("T-cycle experiment") of light and dark.
Skeleton photoperiods mimic the full photoperiods with the light pulses at the times of dawn and dusk only. Although the total time per day that the animal spends in light is quite small, the photoperiodic response is elicited if the night-break pulse (or the second of the two skeleton pulses) falls during the photoinductive phase of the circadian cycle.
However, in skeleton photoperiods, it is often impossible to mimic long photoperiods as the circadian system tends to entrain to the shorter of the two possible interpretations of the skeleton photoperiods. Also, there is often an additional complication of phase-shifting effects of both pulses.
Resonance cycles are light-dark regimes in which the period T (L+D) is a multiple of 12 hours (e.g., LD 6:18, LD 6:30, LD 6:42, LD 6:54, etc.). If the circadian system is involved, every other light pulse will fall onto the photosensitive phase and induce a photoperiodic response in all cycles which are odd multiples of 12 hours (e.g., LD6:18, LD 6:42), while in cycles which are even multiples of 12 hours (e.g., LD 6:30, LD 6:54) the photoperiodic response will not be seen because every light pulse falls only during the insensitive phase of the rhythm of photoperiodic photosensitivity. In resonance cycles, the long periods of darkness may theoretically counter the stimulatory effects of the rare light pulses.
In T-cycle experiments, the period (T) of the LD cycle is different from 24 hours and varies within the limits of entrainment. During entrainment, the phase of the pacemaker is shifted or reset each cycle by an amount (Df) equal to the difference between the freerunning period (t) and the period (T) of the entraining cycle ( t - T = Df). The phase angle (y) between the onset of light and the onset of circadian activity will be characteristic of each T-cycle and can be calculated from the phase-response curve (PRC).
The PRC describes the phase-shifting behavior (lengths of advances or delays) of a circadian system elicited by a specific environmental perturbation (e.g., a pulse of light of defined length, intensity and wavelength) as a function of circadian timing of that perturbation. As a general rule, light pulses given around early subjective night elicit phase-delays, while those given late in the subjective night result in phase-advances.
In LD cycles of different period lengths, the phase angles between the onset of circadian activity and the onset of the light phase of the cycle will be different and predictable from the PRC. Accordingly, on some T-cycles the circadian system will be entrained in such a way that the light pulse illuminates a portion of the photoinducible phase (fi) and result in a measurable physiological photoperiodic response. If the internal coincidence model is correct, the entraining LD cycle will control the phases of different circadian outputs differently, thus changing the phase relationships between two or more circadian rhythms within the organism. Light-dark cycles of certain period lengths (T) will then bring the relevant rhythms into a phase relationship that results in the stimulation of the photoperiodically controlled physiological system.
The T-cycle protocol does not have the same difficulties in interpretation as the other two protocols. Only one light pulse per cycle is used and it has both an entraining and a potential photostimulatory effect. Utilizing different period-length cycles spanning the limits of entrainment of the organism, the light pulse can be brought into every possible phase-relationship with the circadian system. As the animals assume stable entrainment, the light pulse and the circadian system will assume a different phase-angle in each T-cycle. Presumably, in some of the cycles the light will repeatedly illuminate the photoperiodically photosensitive phase leading to the physiological response, while in other T-cycles, the light will be coincident only with the subjective day, leading to the lack of stimulation.
If an internal coincidence mechanism is operating, it is likely that the T-cycles will place the two circadian oscillators in all possible phase-relationships, leading to the stimulation in some and inhibition of response in other values of T. In theory, the two models may have different predictions as to which Ts will be inductive and which not.
So, what's the evidence?
In many organisms short days inhibit while long days stimulate various seasonal responses of physiology and behavior. Various experimental paradigms, including skeleton photoperiods and resonance cycles, demonstrated that it is not the total amount of light, but the precise temporal placing of the light that induces the photoperiodic response, giving the circadian system a role in the measurement and interpretation of daylength.
In a large number of species, including both invertebrates and vertebrates, as well as plants, these protocols have proven the involvement of the circadian system in photoperiodic time measurement, e.g., resonance cycles in male rosefinches, skeleton photoperiods in the rain quail Coturnix coromendelica, T-cycles and resonance cycles in golden hamsters and house finches, etc.
On the other hand, there are some organisms in which the results of all these protocols are not so conclusive. For instance, T-cycle and night-break studies in blackheaded buntings indicate the dependence of the reproductive response on the period (T) of the cycle and on the length of the light pulse within the cycle. In starlings, effects of light intensity, wavelength and pulse length can be seen even in 24 hour cycles.
Although the involvement of the circadian system has been documented in many species, it has not yet been possible to discriminate between the external and the internal coincidence mechanism for photoperiodic time measurement. So far, results of skeleton photoperiod, resonance cycle and T-cycle experiments could be interpreted equally well with both models.
Moreover, in some species it was even difficult to differentiate between the hourglass and circadian models. The vetch aphid (Megoura viciae) is probably the most notorious example. A mathematical model, though, was developed that succesfully describes photoperiodic responses in all of the hundreds of arthropod species that were studied over the past century. The fact that this mathematical model is consistent with all the experimental data on the vetch aphid suggests that it may also use its circadian clock to measure photoperiod, it's just that its clock's putput is of a very small amplitude.
Another species that was difficult to figure out was American chameleon (Anolis carolinensis). Initial studies suggested that hourglass model was operating. However, later work by Linda Hyde utilizing T-cycles showed that the circadian clock is indeed involved in photoperiodic time measurement in this species.
Finally, in Japanese quail (Coturnix japonica), results of skeleton photoperiods were positive, resonance cycles suggest some involvement of the clock but not the complete response, and the T-cycles were negative. This is also a species with a low-amplitude circadian system (i.e., it has a Type 0 PRC), so it is likely that clock is involved but the final demonstration is still waiting for the next creative experimenter to provide.
Category: Clock Tutorials
posted by Bora Zivkovic at 11:54 PM
![]()
![]()






0 Comments:
Post a Comment
<< Home