Does circadian clock regulate clutch-size in birds? A question of appropriatness of the model animal.
The questions of animal models
There are some very good reasons why much of biology is performed in just a handful of model organisms. Techniques get refined and the knowledge can grow incrementally until we can know quite a lot of nitty-gritty details about a lot of bioloigcal processes. One need not start from Square One with every new experiment with every new species. One should, of course, occasionally test how generalizable such findings are to other organisms, but the value of models is hard to dispute.
Which organisms have been chosen as laboratory models often depends on contingencies of history. Somebody at some point in time had a large supply of a particular species, did some good research on it and everything took off from there. Some species proved untractable and were discarded. Others flourished and were adopted by more and more researchers over time. Some models are really good for studying particular areas of biology. The fruitfly is unparalleled as the organism for genetic studies. However, its development is quite unusual so the findings are difficult to generalize even to other Dipterans, even less to other insects, arthropods or other animals.
What makes a good laboratory animal model?
A good animal model should be one about which there is quite a lot of background information. It should be available in large numbers and it should be rather small in size so a lab can keep large numbers of individuals in a relatively small space. Being social also makes housing easier. A good model animal is also easy to keep in captivity, it is easy to feed and easy to handle without too much danger to the technicians. Ideally, it easily breeds in captivity, has a short generation time and large numbers of progeny.
A brief aside about mammalian models
It is not surprising that early studies in anatomy and physiology utilized domesticated animals. Much of 18th and 19th century research was done in dogs, cats, rabbits, sheep, goats, pigs and even horses. Such animals were easily available, there was a lot of existing knowledge about them, and they were easy to house, feed and handle. Furthermore, one could always "sell" such research as useful for advances in agriculture. However, there are definite drawbacks in using these species as well. They tend to be large, with long generation times and with few progeny. Also, being human companions for thousands of years, people get easily attached to them and the anti-vivisectionist movement was quite strong in the 19th century, especially in England.
Thus, after several decades of effort, several new species were domesticated in order to provide the researchers with more tractable and less controversial animals to work with. These include rats, mice, hamsters, guinea pigs, ground squirrels and ferrets. Much of the research in the 20th century was performed on these species. In the last couple of decades, one of those species gained prominence - the mouse - due to its tractability to genetic techniques. Much of the findings from rodent (including mouse) studies have been found to be generalizable to other rodents, to other mammals (including humans) and to other vertebrates. Much has been learned that proved applicable to endangered species and their reproduction in captivity. Rarely is any work done on rodents these days that as its main aim has the better understanding of the rodents. Mainly, the mouse and the rat are the stand-ins for humans and the research is thought of as biomedical research.
Avian models
The bird models have quite a different history. The readily available species are the domesticated birds, better known as poultry - chicken, quail, duck, turkey, goose, pigeon and the guinea hen. There was never a need to move away from these species, as they are small, social, breed fast and a lot, and are viewed as food (and not as "cute pets") thus they were a less likely target of anti-vivisectionists.
Sure, many biologists studied wild species in the field and in the lab. Even today, much work is done in parrots, European starlings, house sparrows, house finches, crows, etc. But wild species are tough to work with. One has to go out and catch a new batch for each new experiment. Poultry is so much easier to handle and breed in the lab.
The lack of a historical "switch" from one group of species to another had one unfortunate consequence. Research on domesticated species of birds is too easily dismissed as Poultry Science, geared towards increased meat and egg production at farms (what's wrong with that? The findings from such research usually make the life for farm birds better, e.g., less stressful). It is sometimes forgotten that many scientists use poultry species with a different motivation altogether - as a model for wild birds, hopefully generalizable even broader to other vertebrates or even all animals. Many findings from poultry research are now being used to help endangered bird species survive and breed in the wild and in captivity.
Why such a long intro?
 Because my lab animal is a domesticated bird, poultry if you wish - the Japanese quail. I will not be unhappy if my research gets picked up by poultry scientists and applied to make the life of farm birds easier, but that is not my main motivation. I am interested in particular aspects of basic science of chronobiology and I found quail to be an excellent model for questions that are difficult or even impossible to do in mammals. I hope that my work is generalizable to wild birds and will help in saving endangered species from extinction. I hope that at least some of my work will be even more broadly generalizable, perhaps to all vertebrates, or even to all of life.
Because my lab animal is a domesticated bird, poultry if you wish - the Japanese quail. I will not be unhappy if my research gets picked up by poultry scientists and applied to make the life of farm birds easier, but that is not my main motivation. I am interested in particular aspects of basic science of chronobiology and I found quail to be an excellent model for questions that are difficult or even impossible to do in mammals. I hope that my work is generalizable to wild birds and will help in saving endangered species from extinction. I hope that at least some of my work will be even more broadly generalizable, perhaps to all vertebrates, or even to all of life.
Japanese quail (Coturnix japonica) has been domesticated about 500 years ago in Japan. The name 'Japanese quail' is used to denote the domesticated birds in laboratories and on farms. Their counterpart in the wild, itself not an endangered species, is usually called Asian Migratory Quail, and it appears that there is not that much difference between wild and domesticated populations in many aspects of their biology. With incubation time of 17 days, maturation time of six weeks, and ability to lay almost an egg a day throughout the year, quail is the mouse of the avian biology lab. The chicken genome has been sequenced. Every gene that has beem looked at is between 95 and 100% identical between the chicken and the quail. It appears that the difference between the two species is mainly due to regulatory regions of the genes driving somewhat different trajectoris of embryonic developments, similarly to the difference between humans and chimps.
An important part of my Masters degree research (but not my PhD) was on the mechanisms by which the circadian system controls reproduction - both seasonally (photoperiodism) and daily (the timing of egg-laying). My MS thesis has been published in these two papers [Note: the first one appears not to be available in its entirety online - you can only see the abstract. I have notified the Journal and once this is fixed I will make the appropriate link to the PDF of the whole paper. Update: The paper is now online and can be accessed here].
Circadian control of egg-laying
Circadian system in Japanese quail, and presumably in many other (perhaps all?) species of birds, is a two-oscillator system. It is as if the brain contains two separate circadian clocks, each with its own properties. These two clocks also interact with each other. One of the clocks is easily and directly entrained by the environmental light-dark cycles. It governs the overt daily rhythms of such things as body temperature, feeding activity, heart rate, blood pressure and release of hormones such as melatonin. The other clock appears not to be directly sensitive to light. It seems that it gets its information about the environmental lighting conditions from its interaction with the first clock. This second clock (or second component of the circadian system) also drives rhythms such as activity and body temperature, but more importantly, it drives the circadian rhythms of ovulation and oviposition. In this species, once an egg is laid, the next follicle ovulates about 45 minutes later. It takes anywhere between 24 and 30 hours for an ovulated egg to be laid. Thus, one egg is laid each day.
The two circadian clocks interact in ways that are possible to describe using models borrowed from the physics of oscillations (remember the pendulum from high school?). One of the predictions from the model is that some biological events can happen only when the two oscillators are in a particular phase-relationship with each other. This phase, in regards to ovulation in birds, is sometimes called "the permissive zone". Only if the ovary is ready to ovulate during this zone, ovulation will occur. Otherwise, it will wait until the next day.
In short daylengths, e.g., during the winter, the permissive zone is so narrow (or even non-existent), so that no ovulation happens at all. The bar on top of the graph indicates the duration of the day - white and night - black. The first line is first day, the line below it is the second day, etc. X-axis is 24 hours long. Bluish area shows the times when the first clock drives body temperature to be above the daily mean and drives the animal to be behaviorally active; the white area shows the times when the first clock determines when the quail is asleep and the melatonin levels are high.
 As the photoperiod increases in spring, the permissive zone appears (between two dotted lines) and is broad enough that ovulation can happen. In relatively short photoperiods (but above the critical photoperiod for the onset of ovulation), the two clocks are quite tightly coupled. They are almost entrained to each other - they exhibit what is called relative coordination, which means that the two clocks can entrain ("lock") to each other for a few cycles but cannot remain phase-locked indefinitely:
As the photoperiod increases in spring, the permissive zone appears (between two dotted lines) and is broad enough that ovulation can happen. In relatively short photoperiods (but above the critical photoperiod for the onset of ovulation), the two clocks are quite tightly coupled. They are almost entrained to each other - they exhibit what is called relative coordination, which means that the two clocks can entrain ("lock") to each other for a few cycles but cannot remain phase-locked indefinitely:
 The first egg gets laid at the beginning of the permissive phase. Each day, the second clock tries to escape from the pull of the first clock and to time the egg-laying a little bit later. Finally, the ovulation reaches the end of the permissive phase and the clutch stops. If everything is OK, prolactin will kick in to prevent the hen from laying any more eggs and she can sit down on her eggs and incubate them. If a predator, like a snake, eats the eggs (or a researcher removes the eggs from the cage) there is no rise in prolactin and the quail starts a new clutch all over again, following the same pattern, after one or two days of pause.
The first egg gets laid at the beginning of the permissive phase. Each day, the second clock tries to escape from the pull of the first clock and to time the egg-laying a little bit later. Finally, the ovulation reaches the end of the permissive phase and the clutch stops. If everything is OK, prolactin will kick in to prevent the hen from laying any more eggs and she can sit down on her eggs and incubate them. If a predator, like a snake, eats the eggs (or a researcher removes the eggs from the cage) there is no rise in prolactin and the quail starts a new clutch all over again, following the same pattern, after one or two days of pause.
A couple of months later, after the bird has succesfully raised a brood of hatchlings, the quail may decide to renest. This is now early summer and the daylength is much longer. The permissive phase is much broader and intrudes even into the night. On the other hand, the coupling between the two clocks is now very weak and the second clock freeruns with its own inherent period which can be anywhere between 26 and 30 hours. The pattern of egg-laying now looks something like this:
 Now compare the last two figures. One thing that you will notice is that the clutch consisted of more eggs in shorter photoperiod (spring), than in the long photoperiod (summer). This is something that has been observed in a number of species of birds in the wild. The above figures are schematics. Here are some real data. Every 10-minute period that is black denotes the time when body temperature is above the daily mean. The light bars on top show when the lighs were on (white) and off (black). Circles show times when eggs were laid. On the top is a quail in LD 14:10. In the middle is a quail in LD 18:6. On the bottom are two examples of oviposition patterns during a transition from LD 14:10 to LD 18:6 (all from [1]):
Now compare the last two figures. One thing that you will notice is that the clutch consisted of more eggs in shorter photoperiod (spring), than in the long photoperiod (summer). This is something that has been observed in a number of species of birds in the wild. The above figures are schematics. Here are some real data. Every 10-minute period that is black denotes the time when body temperature is above the daily mean. The light bars on top show when the lighs were on (white) and off (black). Circles show times when eggs were laid. On the top is a quail in LD 14:10. In the middle is a quail in LD 18:6. On the bottom are two examples of oviposition patterns during a transition from LD 14:10 to LD 18:6 (all from [1]):


 Not so obvious, but the time-difference between two successive eggs in spring is just a little bit longer than 24 hours. In summer, this difference is 26-30 hours. This means that in summer, each follicle has more time to get filled by filtering of blood and each egg has more time to spend in the oviduct and the shell-gland. One should expect that summer eggs will be, on average, larger, heavier and with thicker shell than eggs laid in early spring. Evolutionary theory predicts the existence of a trade-off between egg-number (i.e., clutch-size) and egg-size. It is almost like a switch between r-strategy and K-strategy. In spring, premium is put on quantity and in summer on quality of progeny. The quail literature suggests that the circadian clock is the physiological mechanism underlying this switch in evolutionary strategies.
Not so obvious, but the time-difference between two successive eggs in spring is just a little bit longer than 24 hours. In summer, this difference is 26-30 hours. This means that in summer, each follicle has more time to get filled by filtering of blood and each egg has more time to spend in the oviduct and the shell-gland. One should expect that summer eggs will be, on average, larger, heavier and with thicker shell than eggs laid in early spring. Evolutionary theory predicts the existence of a trade-off between egg-number (i.e., clutch-size) and egg-size. It is almost like a switch between r-strategy and K-strategy. In spring, premium is put on quantity and in summer on quality of progeny. The quail literature suggests that the circadian clock is the physiological mechanism underlying this switch in evolutionary strategies.
How generalizable is this finding?
Apart from Japanese quail, few other species have been studied. The sister species, European Migratory Quail (Coturnix coturnix), which is not domesticated, has virtually identical egg-laying patterns. In domestic chickens, it looks very similar, except that the first egg is laid at dawn (as opposed to noon) and the last egg is laid around noon (as opposed to evening). A couple of studies in turkeys also suggest the same mechanism.
Can we generalize this finding to all Gallinaceous birds at least? Sounds reasonable, but I do not know. Is the same mechanism operating in birds from other Orders? Hummingbirds? Owls? Ostriches?
There is an interaction between estrous cycle and circadian cycle in rodents. Sex steroid hormones that are released during ovulation have effects on circadian clocks in several species of birds and mammals (presumably also humans). Does this information make it seem more likely that the quail data are generalizable to all birds?
Every time I ask a friend who studies wild birds at what times the eggs are laid, the answer is "I don't know - we usually find the eggs in the morning". This means that the egg could have been laid at any time between 5pm and 9am - a very broad region. No temporal patterns appear to be known in wild birds. So, if you study wild birds, please let me know at what time of day the eggs are laid. Let's see if the data gathered from poultry are useful for the study of wild species as well.
[1] Bora D. Zivkovic, Herbert Underwood, and Thomas Siopes, Circadian Ovulatory Rhythms in Japanese Quail: Role of Ocular and Extraocular Pacemakers, JOURNAL OF BIOLOGICAL RHYTHMS, Vol. 15 No. 2, April 2000 172-183
There are some very good reasons why much of biology is performed in just a handful of model organisms. Techniques get refined and the knowledge can grow incrementally until we can know quite a lot of nitty-gritty details about a lot of bioloigcal processes. One need not start from Square One with every new experiment with every new species. One should, of course, occasionally test how generalizable such findings are to other organisms, but the value of models is hard to dispute.
Which organisms have been chosen as laboratory models often depends on contingencies of history. Somebody at some point in time had a large supply of a particular species, did some good research on it and everything took off from there. Some species proved untractable and were discarded. Others flourished and were adopted by more and more researchers over time. Some models are really good for studying particular areas of biology. The fruitfly is unparalleled as the organism for genetic studies. However, its development is quite unusual so the findings are difficult to generalize even to other Dipterans, even less to other insects, arthropods or other animals.
What makes a good laboratory animal model?
A good animal model should be one about which there is quite a lot of background information. It should be available in large numbers and it should be rather small in size so a lab can keep large numbers of individuals in a relatively small space. Being social also makes housing easier. A good model animal is also easy to keep in captivity, it is easy to feed and easy to handle without too much danger to the technicians. Ideally, it easily breeds in captivity, has a short generation time and large numbers of progeny.
A brief aside about mammalian models
It is not surprising that early studies in anatomy and physiology utilized domesticated animals. Much of 18th and 19th century research was done in dogs, cats, rabbits, sheep, goats, pigs and even horses. Such animals were easily available, there was a lot of existing knowledge about them, and they were easy to house, feed and handle. Furthermore, one could always "sell" such research as useful for advances in agriculture. However, there are definite drawbacks in using these species as well. They tend to be large, with long generation times and with few progeny. Also, being human companions for thousands of years, people get easily attached to them and the anti-vivisectionist movement was quite strong in the 19th century, especially in England.
Thus, after several decades of effort, several new species were domesticated in order to provide the researchers with more tractable and less controversial animals to work with. These include rats, mice, hamsters, guinea pigs, ground squirrels and ferrets. Much of the research in the 20th century was performed on these species. In the last couple of decades, one of those species gained prominence - the mouse - due to its tractability to genetic techniques. Much of the findings from rodent (including mouse) studies have been found to be generalizable to other rodents, to other mammals (including humans) and to other vertebrates. Much has been learned that proved applicable to endangered species and their reproduction in captivity. Rarely is any work done on rodents these days that as its main aim has the better understanding of the rodents. Mainly, the mouse and the rat are the stand-ins for humans and the research is thought of as biomedical research.
Avian models
The bird models have quite a different history. The readily available species are the domesticated birds, better known as poultry - chicken, quail, duck, turkey, goose, pigeon and the guinea hen. There was never a need to move away from these species, as they are small, social, breed fast and a lot, and are viewed as food (and not as "cute pets") thus they were a less likely target of anti-vivisectionists.
Sure, many biologists studied wild species in the field and in the lab. Even today, much work is done in parrots, European starlings, house sparrows, house finches, crows, etc. But wild species are tough to work with. One has to go out and catch a new batch for each new experiment. Poultry is so much easier to handle and breed in the lab.
The lack of a historical "switch" from one group of species to another had one unfortunate consequence. Research on domesticated species of birds is too easily dismissed as Poultry Science, geared towards increased meat and egg production at farms (what's wrong with that? The findings from such research usually make the life for farm birds better, e.g., less stressful). It is sometimes forgotten that many scientists use poultry species with a different motivation altogether - as a model for wild birds, hopefully generalizable even broader to other vertebrates or even all animals. Many findings from poultry research are now being used to help endangered bird species survive and breed in the wild and in captivity.
Why such a long intro?
 Because my lab animal is a domesticated bird, poultry if you wish - the Japanese quail. I will not be unhappy if my research gets picked up by poultry scientists and applied to make the life of farm birds easier, but that is not my main motivation. I am interested in particular aspects of basic science of chronobiology and I found quail to be an excellent model for questions that are difficult or even impossible to do in mammals. I hope that my work is generalizable to wild birds and will help in saving endangered species from extinction. I hope that at least some of my work will be even more broadly generalizable, perhaps to all vertebrates, or even to all of life.
Because my lab animal is a domesticated bird, poultry if you wish - the Japanese quail. I will not be unhappy if my research gets picked up by poultry scientists and applied to make the life of farm birds easier, but that is not my main motivation. I am interested in particular aspects of basic science of chronobiology and I found quail to be an excellent model for questions that are difficult or even impossible to do in mammals. I hope that my work is generalizable to wild birds and will help in saving endangered species from extinction. I hope that at least some of my work will be even more broadly generalizable, perhaps to all vertebrates, or even to all of life.Japanese quail (Coturnix japonica) has been domesticated about 500 years ago in Japan. The name 'Japanese quail' is used to denote the domesticated birds in laboratories and on farms. Their counterpart in the wild, itself not an endangered species, is usually called Asian Migratory Quail, and it appears that there is not that much difference between wild and domesticated populations in many aspects of their biology. With incubation time of 17 days, maturation time of six weeks, and ability to lay almost an egg a day throughout the year, quail is the mouse of the avian biology lab. The chicken genome has been sequenced. Every gene that has beem looked at is between 95 and 100% identical between the chicken and the quail. It appears that the difference between the two species is mainly due to regulatory regions of the genes driving somewhat different trajectoris of embryonic developments, similarly to the difference between humans and chimps.
An important part of my Masters degree research (but not my PhD) was on the mechanisms by which the circadian system controls reproduction - both seasonally (photoperiodism) and daily (the timing of egg-laying). My MS thesis has been published in these two papers [Note: the first one appears not to be available in its entirety online - you can only see the abstract. I have notified the Journal and once this is fixed I will make the appropriate link to the PDF of the whole paper. Update: The paper is now online and can be accessed here].
Circadian control of egg-laying
Circadian system in Japanese quail, and presumably in many other (perhaps all?) species of birds, is a two-oscillator system. It is as if the brain contains two separate circadian clocks, each with its own properties. These two clocks also interact with each other. One of the clocks is easily and directly entrained by the environmental light-dark cycles. It governs the overt daily rhythms of such things as body temperature, feeding activity, heart rate, blood pressure and release of hormones such as melatonin. The other clock appears not to be directly sensitive to light. It seems that it gets its information about the environmental lighting conditions from its interaction with the first clock. This second clock (or second component of the circadian system) also drives rhythms such as activity and body temperature, but more importantly, it drives the circadian rhythms of ovulation and oviposition. In this species, once an egg is laid, the next follicle ovulates about 45 minutes later. It takes anywhere between 24 and 30 hours for an ovulated egg to be laid. Thus, one egg is laid each day.
The two circadian clocks interact in ways that are possible to describe using models borrowed from the physics of oscillations (remember the pendulum from high school?). One of the predictions from the model is that some biological events can happen only when the two oscillators are in a particular phase-relationship with each other. This phase, in regards to ovulation in birds, is sometimes called "the permissive zone". Only if the ovary is ready to ovulate during this zone, ovulation will occur. Otherwise, it will wait until the next day.
In short daylengths, e.g., during the winter, the permissive zone is so narrow (or even non-existent), so that no ovulation happens at all. The bar on top of the graph indicates the duration of the day - white and night - black. The first line is first day, the line below it is the second day, etc. X-axis is 24 hours long. Bluish area shows the times when the first clock drives body temperature to be above the daily mean and drives the animal to be behaviorally active; the white area shows the times when the first clock determines when the quail is asleep and the melatonin levels are high.
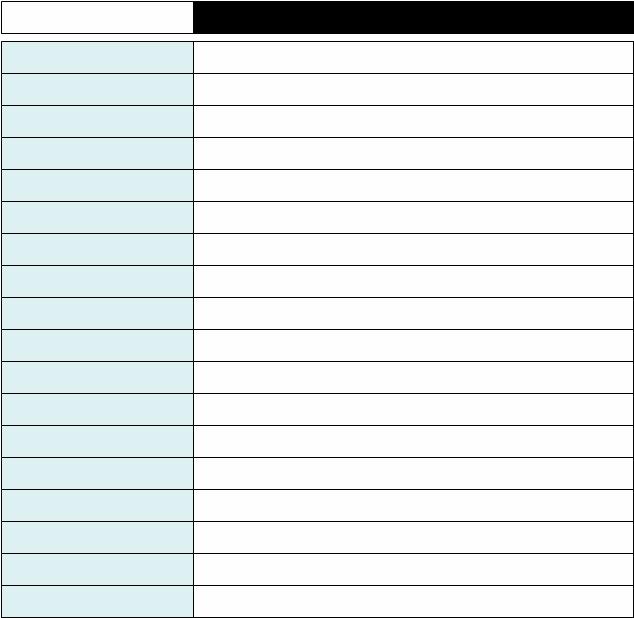 As the photoperiod increases in spring, the permissive zone appears (between two dotted lines) and is broad enough that ovulation can happen. In relatively short photoperiods (but above the critical photoperiod for the onset of ovulation), the two clocks are quite tightly coupled. They are almost entrained to each other - they exhibit what is called relative coordination, which means that the two clocks can entrain ("lock") to each other for a few cycles but cannot remain phase-locked indefinitely:
As the photoperiod increases in spring, the permissive zone appears (between two dotted lines) and is broad enough that ovulation can happen. In relatively short photoperiods (but above the critical photoperiod for the onset of ovulation), the two clocks are quite tightly coupled. They are almost entrained to each other - they exhibit what is called relative coordination, which means that the two clocks can entrain ("lock") to each other for a few cycles but cannot remain phase-locked indefinitely: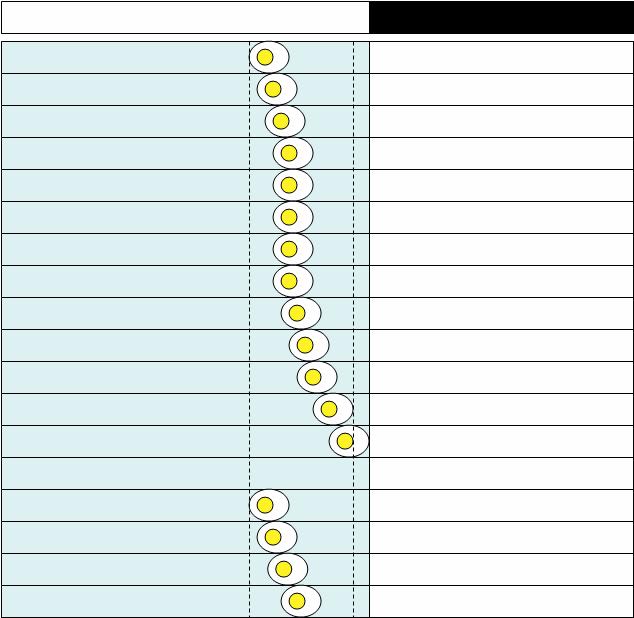 The first egg gets laid at the beginning of the permissive phase. Each day, the second clock tries to escape from the pull of the first clock and to time the egg-laying a little bit later. Finally, the ovulation reaches the end of the permissive phase and the clutch stops. If everything is OK, prolactin will kick in to prevent the hen from laying any more eggs and she can sit down on her eggs and incubate them. If a predator, like a snake, eats the eggs (or a researcher removes the eggs from the cage) there is no rise in prolactin and the quail starts a new clutch all over again, following the same pattern, after one or two days of pause.
The first egg gets laid at the beginning of the permissive phase. Each day, the second clock tries to escape from the pull of the first clock and to time the egg-laying a little bit later. Finally, the ovulation reaches the end of the permissive phase and the clutch stops. If everything is OK, prolactin will kick in to prevent the hen from laying any more eggs and she can sit down on her eggs and incubate them. If a predator, like a snake, eats the eggs (or a researcher removes the eggs from the cage) there is no rise in prolactin and the quail starts a new clutch all over again, following the same pattern, after one or two days of pause.A couple of months later, after the bird has succesfully raised a brood of hatchlings, the quail may decide to renest. This is now early summer and the daylength is much longer. The permissive phase is much broader and intrudes even into the night. On the other hand, the coupling between the two clocks is now very weak and the second clock freeruns with its own inherent period which can be anywhere between 26 and 30 hours. The pattern of egg-laying now looks something like this:
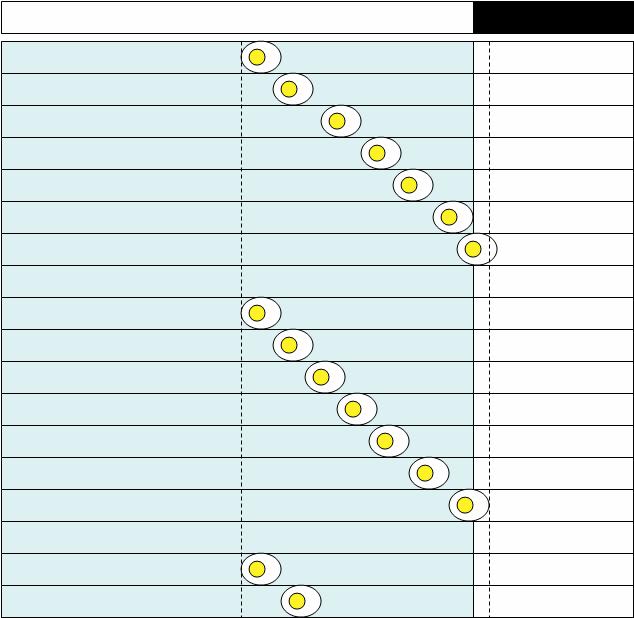 Now compare the last two figures. One thing that you will notice is that the clutch consisted of more eggs in shorter photoperiod (spring), than in the long photoperiod (summer). This is something that has been observed in a number of species of birds in the wild. The above figures are schematics. Here are some real data. Every 10-minute period that is black denotes the time when body temperature is above the daily mean. The light bars on top show when the lighs were on (white) and off (black). Circles show times when eggs were laid. On the top is a quail in LD 14:10. In the middle is a quail in LD 18:6. On the bottom are two examples of oviposition patterns during a transition from LD 14:10 to LD 18:6 (all from [1]):
Now compare the last two figures. One thing that you will notice is that the clutch consisted of more eggs in shorter photoperiod (spring), than in the long photoperiod (summer). This is something that has been observed in a number of species of birds in the wild. The above figures are schematics. Here are some real data. Every 10-minute period that is black denotes the time when body temperature is above the daily mean. The light bars on top show when the lighs were on (white) and off (black). Circles show times when eggs were laid. On the top is a quail in LD 14:10. In the middle is a quail in LD 18:6. On the bottom are two examples of oviposition patterns during a transition from LD 14:10 to LD 18:6 (all from [1]):

How generalizable is this finding?
Apart from Japanese quail, few other species have been studied. The sister species, European Migratory Quail (Coturnix coturnix), which is not domesticated, has virtually identical egg-laying patterns. In domestic chickens, it looks very similar, except that the first egg is laid at dawn (as opposed to noon) and the last egg is laid around noon (as opposed to evening). A couple of studies in turkeys also suggest the same mechanism.
Can we generalize this finding to all Gallinaceous birds at least? Sounds reasonable, but I do not know. Is the same mechanism operating in birds from other Orders? Hummingbirds? Owls? Ostriches?
There is an interaction between estrous cycle and circadian cycle in rodents. Sex steroid hormones that are released during ovulation have effects on circadian clocks in several species of birds and mammals (presumably also humans). Does this information make it seem more likely that the quail data are generalizable to all birds?
Every time I ask a friend who studies wild birds at what times the eggs are laid, the answer is "I don't know - we usually find the eggs in the morning". This means that the egg could have been laid at any time between 5pm and 9am - a very broad region. No temporal patterns appear to be known in wild birds. So, if you study wild birds, please let me know at what time of day the eggs are laid. Let's see if the data gathered from poultry are useful for the study of wild species as well.
[1] Bora D. Zivkovic, Herbert Underwood, and Thomas Siopes, Circadian Ovulatory Rhythms in Japanese Quail: Role of Ocular and Extraocular Pacemakers, JOURNAL OF BIOLOGICAL RHYTHMS, Vol. 15 No. 2, April 2000 172-183
posted by Bora Zivkovic at 1:43 AM
![]()
![]()






0 Comments:
Post a Comment
<< Home