Clocks in Bacteria II: Adaptive Function of Clocks in Cyanobacteria
In the previous two posts, here and here, I have mentioned how the discovery of circadian clocks in Cyanobacteria changed the way we think about the origin and evolution of circadian clocks. Quite soon after the initial discovery, the team from Carl Johnson's laboratory published two papers [1,2] describing a more direct test of adaptive function of circadian clocks in the Synechococcus elongatus.
 Wild-type and various clock-mutants in Synechoccocus, when raised in isolation in light-dark cycles, have comparable reproductive rates. When raised in constant light, they fare even a little better, i.e., multiply faster. Thus, in isolation, clock does not appear to confer adaptive advantage.
Wild-type and various clock-mutants in Synechoccocus, when raised in isolation in light-dark cycles, have comparable reproductive rates. When raised in constant light, they fare even a little better, i.e., multiply faster. Thus, in isolation, clock does not appear to confer adaptive advantage.
 However, when the strains are cultured together, two strains grown in the same petri-dish, and exposed to light-dark cycle, the strain whose endogenous period is closer to the period of the environmental cycle "wins" the contest. This suggests that circadian clock confers fitness in rhythmic environments. In constant light, arrhythmic mutants outperform rhythmic strains.
However, when the strains are cultured together, two strains grown in the same petri-dish, and exposed to light-dark cycle, the strain whose endogenous period is closer to the period of the environmental cycle "wins" the contest. This suggests that circadian clock confers fitness in rhythmic environments. In constant light, arrhythmic mutants outperform rhythmic strains.
 Here is how Johnson describes the experiments (from a book chapter not available online):
Here is how Johnson describes the experiments (from a book chapter not available online):
 Here is Johnson [3] again:
Here is Johnson [3] again:
In his book Adaptation and Environment (1990), Robert Brandon came up with five criteria that need to be satisfied in order to determine if a trait is an adaptation (thanks to Robert Skipper for a reminder of this):
 The early cyanobacterial studies have shown criterion #3 to be correct. The competitive assay studies started cracking the criterion #2. In the next post on this topic, I will describe some studies that started investigating the criterion #5, with some additional evidence for criteria Nos. 1, 2 and 4. Apparently, we still have a long way to go. Johnson again:
The early cyanobacterial studies have shown criterion #3 to be correct. The competitive assay studies started cracking the criterion #2. In the next post on this topic, I will describe some studies that started investigating the criterion #5, with some additional evidence for criteria Nos. 1, 2 and 4. Apparently, we still have a long way to go. Johnson again:
[1] YAN OUYANG, CAROL R. ANDERSSON, TAKAO KONDO, SUSAN S. GOLDEN, AND CARL HIRSCHIE JOHNSON, Resonating circadian clocks enhance fitness in cyanobacteria, Proc. Natl. Acad. Sci. USA, Vol. 95, pp. 8660–8664, July 1998
[2] Mark A. Woelfle, Yan Ouyang, Kittiporn Phanvijhitsiri and Carl Hirschie Johnson, The Adaptive Value of Circadian Clocks: An Experimental Assessment in Cyanobacteria, Current Biology, Vol. 14, 1481–1486, August 24, 2004,
[3] Carl Hirschie Johnson, Testing the Adaptive Value of Circadian Systems, Methods in Enzymology, Volume 393 , 2005, Pages 818-837
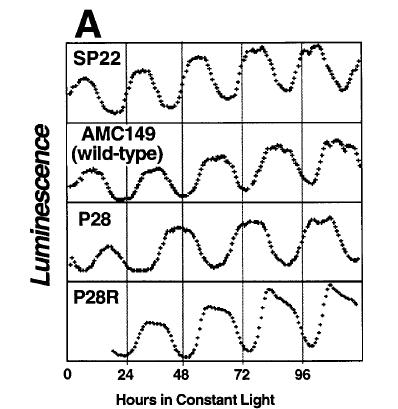 Wild-type and various clock-mutants in Synechoccocus, when raised in isolation in light-dark cycles, have comparable reproductive rates. When raised in constant light, they fare even a little better, i.e., multiply faster. Thus, in isolation, clock does not appear to confer adaptive advantage.
Wild-type and various clock-mutants in Synechoccocus, when raised in isolation in light-dark cycles, have comparable reproductive rates. When raised in constant light, they fare even a little better, i.e., multiply faster. Thus, in isolation, clock does not appear to confer adaptive advantage. However, when the strains are cultured together, two strains grown in the same petri-dish, and exposed to light-dark cycle, the strain whose endogenous period is closer to the period of the environmental cycle "wins" the contest. This suggests that circadian clock confers fitness in rhythmic environments. In constant light, arrhythmic mutants outperform rhythmic strains.
However, when the strains are cultured together, two strains grown in the same petri-dish, and exposed to light-dark cycle, the strain whose endogenous period is closer to the period of the environmental cycle "wins" the contest. This suggests that circadian clock confers fitness in rhythmic environments. In constant light, arrhythmic mutants outperform rhythmic strains.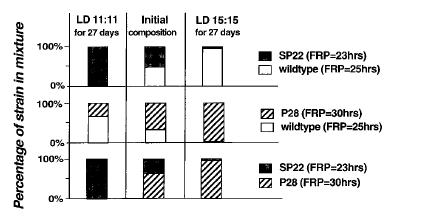 Here is how Johnson describes the experiments (from a book chapter not available online):
Here is how Johnson describes the experiments (from a book chapter not available online):"The authors’ laboratory tested the adaptive significance of circadian programs by using competition experiments between different strains of the cyanobacterium Synechococcus elongatus (Ouyang et al., 1998; Woelfle et al., 2004). For asexual microbes such as S. elongatus, differential growth of one strain under competition with other strains is a good measure of reproductive fitness. In pure culture, because the strains grew at about the same rate in constant light and in LD cycles, there did not appear to be a significant advantage or disadvantage in having different circadian periods when the strains were grown individually. The fitness test was to mix different strains together and to grow them in competition to determine whether the composition of the population changes as a function of time. The cultures were diluted at intervals to allow growth to continue. Different period mutants were used to answer the question, ‘‘Does having a period that is similar to the period of the environmental cycle enhance fitness?’’ The circadian phenotypes of the strains used had freerunning periods of about 22 h (B22a, C22a) and 30 h (A30a, C28a). These strains were determined by point mutations in three different clock genes: kaiA (A30a), kaiB (B22a), and kaiC (C22a, C28a). Wild type has a period of about 25 h under these conditions. When each of the strains was mixed with another strain and grown together in competition, a pattern emerged that depended on the frequency of the LD cycle and the circadian period. When grown on a 22-h cycle (LD 11:11), the 22 h-period mutants could overtake wild type in the mixed cultures. On a 30-h cycle (LD 15:15), the 30 h-period mutants could defeat either wild type or the 22 h-period mutants. On a ‘‘normal’’ 24-h cycle (LD 12:12), the wild-type strain could overgrow either mutant (Ouyang et al., 1998). Note that over many cycles, each of these LD conditions have equal amounts of light and dark (which is important, as photosynthetic cyanobacteria derive their energy from light); it is only the frequency of light versus dark that differs among the LD cycles. Figure 1 shows results from the competition between wild type and the mutant strains (Ouyang et al., 1998).It is telling how many control experiments they had to do in order to eliminate various alternative explanations. They had to show that mutations in clock genes do not have additional effects on the ability of the cell to grow and reproduce. Check. They had to show that clock mutations do not affect the ability of the cells to utilize the food and light energy. Check. They had to show that clock mutations do not affect any conceivable way by which one strain can, perhaps by secreting chemicals, actively disrupt the health of the other strain. Check. And in the end, although they demonstrated that "resonance", i.e., similarity between environmental cycle and the intrinsic period confers some advantage, they still could not state with certainty that this "proves" that the circadian clock has an adaptive function.
Clearly, the strain whose period most closely matched that of the LD cycle eliminated the competitor. Under a nonselective condition (in this case, constant light), each strain was able to maintain itself in the mixed cultures. Because the mutant strains could defeat the wild-type strain in LD cycles in which the periods are similar to their endogenous periods, the differential effects that were observed are likely to result from the differences in the circadian clock. A genetic test was also performed to demonstrate that the clock gene mutation was specifically responsible for the differential effects in the competition experiment (Ouyang et al., 1998). Because the growth rate of the various cyanobacterial strains in pure culture is not detectably different, these results are most likely an example of ‘‘soft selection’’ where the reduced fitness of one genotype is seen only under competition (Futuyma, 1998).
In a test of the extrinsic versus intrinsic value of the clock system of cyanobacteria, wild type was competed with an apparently arrhythmic strain (CLAb). As shown in Fig. 2, the arrhythmic strain was defeated rapidly by wild type in LD 12:12, but under competition in constant light, the arrhythmic strain grew slightly better than wild type (Woelfle et al., 2004). Taken together, results show that an intact clock system whose freerunning period is consonant with the environment significantly enhances the reproductive fitness of cyanobacteria in rhythmic environments; however, this same clock system provides no adaptive advantage in constant environments and may even be slightly detrimental to this organism. Therefore, the clock system does not appear to confer an intrinsic value for cyanobacteria in constant conditions."
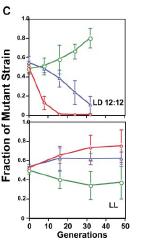 Here is Johnson [3] again:
Here is Johnson [3] again:"The original adaptation of circadian clocks was presumably to enhance reproductive fitness in natural environments, which are cyclic (24h) conditions. We can refer to this situation as an adaptation to extrinsic conditions. However, some researchers have proposed that circadian clocks may additionally provide an "intrinsic" adaptive value (Klarsfeld 1998; Paranjpe 2003 and Pittendrigh 1993). That is, circadian pacemakers may have evolved to become an intrinsic part of internal temporal organization and, as such, may have become intertwined with other traits that influence reproductive fitness in addition to their original role for adaptation to environmental cycles. Note that a rigorous evolutionary biologist would no longer consider an intrinsic value for clocks to be an adaptation if their original extrinsic value has been lost. However, if clocks retain extrinsic value and additionally accrue intrinsic value, then they would still be considered an adaptation."Testing if a trait is an adaptation is a very difficult task. Testing if something as ubiquitous as a circadian clock is an adaptation is even harder. Can you imagine testing if using ATP for energy storage, or using DNA for information storage are adaptations? Are there organisms that do not use these, so we can use them in comparative or competitive studies?
In his book Adaptation and Environment (1990), Robert Brandon came up with five criteria that need to be satisfied in order to determine if a trait is an adaptation (thanks to Robert Skipper for a reminder of this):
"One must have
1. evidence that selection has occurred;
2. an ecological explanation of the fact that some types are better adapted than others;
3. evidence that the trait in question is heritable;
4. information about the structure of the population, including both demic structure and the structure of the selective environment;
5. phylogenetic information concerning what has evolved from what."
 The early cyanobacterial studies have shown criterion #3 to be correct. The competitive assay studies started cracking the criterion #2. In the next post on this topic, I will describe some studies that started investigating the criterion #5, with some additional evidence for criteria Nos. 1, 2 and 4. Apparently, we still have a long way to go. Johnson again:
The early cyanobacterial studies have shown criterion #3 to be correct. The competitive assay studies started cracking the criterion #2. In the next post on this topic, I will describe some studies that started investigating the criterion #5, with some additional evidence for criteria Nos. 1, 2 and 4. Apparently, we still have a long way to go. Johnson again:An example from the circadian literature of a ‘‘just-so’’ story that the author has personally promulgated is that of ‘‘temporal separation’’ of photosynthesis and nitrogen fixation in cyanobacteria. In nitrogen-fixing unicellular bacteria, nitrogen fixation is often phased to occur at night. Nitrogen (N2) fixation is inhibited by low levels of oxygen, which poses a dilemma for photosynthetic bacteria because photosynthesis generates oxygen. Mitsui et al. (1986) proposed that the nocturnal phasing of nitrogen fixation was an adaptation to permit N2 fixation to occur when photosynthesis was not evolving oxygen, and the author has repeated this hypothesis in several publications (Johnson et al., 1996, 1998). This hypothesis would predict that cyanobacterial growth in constant light would be slower than in a light/dark (LD) cycle because nitrogen fixation would be inhibited under these conditions and therefore the growing cells might rapidly become starved for metabolically available nitrogen. The problem is that cyanobacteria grow perfectly well in constant light—in fact, they grow faster in constant light than in LD cycles, presumably because of the extra energy they derive from the additional photosynthesis. This result is inconsistent with the ‘‘temporal separation’’ hypothesis. It does not mean that the ‘‘temporal separation’’ hypothesis is incorrect—in fact, the author believes that under appropriate (but as yet untested) conditions of medium, light, and carbon dioxide, the ‘‘temporal separation’’ hypothesis will emerge triumphant. Nevertheless, the point here is that ‘‘temporal separation in cyanobacteria’’ is an example of a ‘‘just-so’’ circadian story that we like to tell without its being rigorously supported by appropriate data. This was the conclusion of Gould and Lewontin (1979) for many investigations in the field of population biology, and this criticism is on target."
[1] YAN OUYANG, CAROL R. ANDERSSON, TAKAO KONDO, SUSAN S. GOLDEN, AND CARL HIRSCHIE JOHNSON, Resonating circadian clocks enhance fitness in cyanobacteria, Proc. Natl. Acad. Sci. USA, Vol. 95, pp. 8660–8664, July 1998
[2] Mark A. Woelfle, Yan Ouyang, Kittiporn Phanvijhitsiri and Carl Hirschie Johnson, The Adaptive Value of Circadian Clocks: An Experimental Assessment in Cyanobacteria, Current Biology, Vol. 14, 1481–1486, August 24, 2004,
[3] Carl Hirschie Johnson, Testing the Adaptive Value of Circadian Systems, Methods in Enzymology, Volume 393 , 2005, Pages 818-837
posted by Bora Zivkovic at 9:09 AM
![]()
![]()






1 Comments:
Heterocysts - next week! Nostoc and such. Synechococcus is not.
Post a Comment
<< Home