If you know what Chossat's Effect is, I guess you are a) a physiologist, b) expert in thermoregulation, and c) old. This is term that got expunged from the scientific lexicon a few decades ago, in an effort - correct me if I am wrong on this - spearheaded by the U.S.
textbook companies, to replace scientific terminology named after the discoverers (and sometimes even Latin and Greek terms) with bland English neologisms.
But I love Schwann's Cells, Fallopian Tubes (or Mullerian Ducts), Purkinje Fibers, Broca's Area and the amazing Bundle of His! Those terms are memorable, make it easy to sneak in some historical context into teaching science, and have an emotional effect of bringing forth images of ancient scientists working under candlelight, sacrificing their eyesight and health, their social standing and sometimes even their lives, in the feverish hunger for knowledge.
So, what is Chossat's Effect? It comes from a certain 19th century French scientist who was studying the physiology of starvation [1]. The 'modern' term for this effect is "fasting-induced nocturnal hypothernia" (doesn't that sound like something that would prompt the students in the classroom to immediatelly stop paying attention to the teacher and instead pick-up their cell-phones and start text-messaging their friends?).
Actually, this is a very interesting area of research that is very tightly connected to circadian biology. This post is likely to be long, so feel free to skim and just focus on the first part if you are into birds, second part if you are interested in mammals, and the last part if you are into humans.
BirdsAll warm-blooded animals (and yes, that includes at least some reptiles, not to mention a few heat-producing plants like stink-cabbage) exhibit a daily rhythm of body temperature. If an animal is active during the day (diurnal) and sleeps during the night, reducing the metabolic rate during the night is a good way to save energy.
Some of the smallest birds, like swifts and hummingbirds, need to feed continuously in order to stay alive. At night, when they are not able to forage (flowers are closed, it's hard to see, and owls are hunting at the time), they drop their metabolic rate, and thus body temperature, quite dramatically. The body temperature gets down as low as the environmental temperature, sometimes daringly close to the freezing point. The total drop can be as large as 40 degrees Celsius in some instances! This is called
daily torpor (yup, click on that link - it is an excellent blog post) and the metabolic rate drops as much as 95% [2, 3]. This is like full-scale winter hibernation EVERY DAY!
Chossat's effect does not refer to daily torpor, though. It describes a drop in temperature during the night that is larger than the usual circadian fluctuation, in animals undergoing fasting, e.g., during spells of very bad weather (e.g., hurricanes).
Normal amplitude (daily maximum minus nightly minimum) of body temperature in birds with normal access to food ranges between about 1 and 2 degrees Celsius. For instance, a daily maximum may be 41 degrees and the nightly minimum may be 39 degrees (yes, the birds are much warmer than mammals, which makes them inhospitable to microbes that cause many mammalian diseases), which calculates to 2 degrees of amplitude.
During fasting (or food deprivation in the laboratory), the nightly minima drop down to lower levels than in fed birds. The minimum gets lower and lower with each additional night. Importantly, the daily maxima do not change at all. It is thought that it is advantageous for birds to retain their normal metabolic rates during the day so they can immediately resume foraging once the bad weather subsides. Also, if the bad weather persists for too long, the birds need the daytime metabolic rates in order to fly away [4].
According to John Wingfield's "Emergency Life-History Stage" hypothesis [5], an individual's perception of inclement weather directly affect the levels of stress hormones (e.g., corticosterone). An individual who does not perceive the bad weather to be "too bad", will reduce daytime activity and reduce night-time temperature in order to save energy - this individual has made a decision to sit it out.
On the other hand, an individual who perceives bad weather to be "really bad" (or if it lasts too long) will have higher levels of stress hormones and will attempt to fly away during the day. This is not the same mechanism as the seasonal migration, which is usually a nocturnal flight, i.e., they do not experience
Zugunruhe, just stress. Stressed birds do not attempt to escape at night, at which time they have allowed their body temperature to drop by several degrees.
Nocturnal hypothermia has been studied in a large number of species of birds (see, for examples, references # 6-12), but most of the work was performed on pigeons [13-15] and quail [16]. Not all avian species exhibit this response. Laurilla at al. [18] write:
"On the other hand, many large birds that are adapted to long fasting periods as a part of their life histories, e.g. penguins and geese (Cherel et al., 1988; Castellini andRea, 1992), owls (Hohtola et al., 1994) and some raptors (McKechnie andLovegrove, 1999) do not show marked hypothermia during fasting. Some species enter hypothermia upon food restriction only when isolated from conspecifics in a laboratory environment, while in the field they remain normothermic by huddling. These observations have even led some authors to question the usefulness of the concept of hypothermia (Lovegrove andSmith, 2003)."[8]

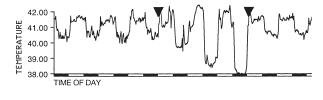
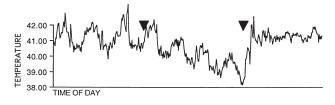
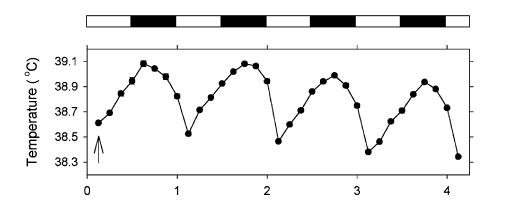
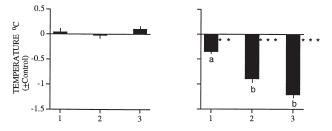
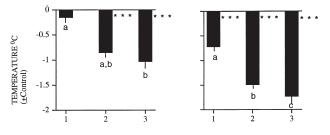
Here is a graphic example of a fasting-induced nocturnal hypothermia in quail (from[17]). The period between the two triangles is the time (3 days) during which the birds had water but no food. Before and after, birds were fed
ad libitum. Below is a graph that shows the difference between the temperature minima during the first, second and third day (left) and night (right) of food deprivation in comparison to the last three days and nights of normal feeding prior to the fasting treatment:
Much of the
more recent research is looking at other environmental cues that can modify the Chossat's effect, as well as the involvement of the circadian clock in this time-specific form of

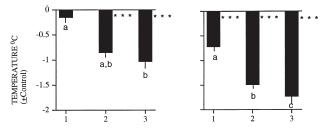
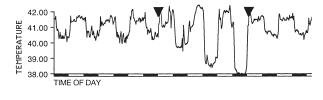
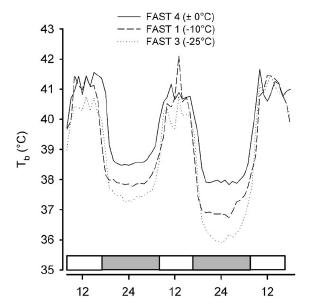
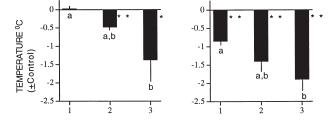
thermoreguluation. For instance, some of the ambient cues that affect the response include ambient temperature [16, 20], ambient light [17], photoperiod [18, 19], single vs. repeated fasting [18, 19], caloric food restriction vs. complete food deprivation [13], social situation, e.g., opportunity for huddling [8] and presence of stationary vs. flying predators [19, 20]. Here is an example of an effect of ambient temperature on nocturnal hypothermia in fasted pigeons (from [20]). Lower the ambient temperature, deepeer the Chossat's effect:
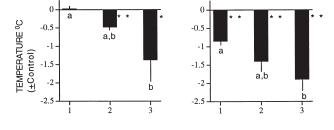
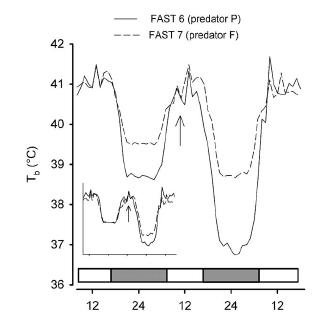
Here is the effect of the presence of a predator (from [2]). In the presence of a perched hawk (P), nocturnal hypothermia reached normally low levels. In the presence of the flying hawk (F), temperature did not drop as much. Presumably, the pigeons kept the metabolic rate high enough to be able to fly fast if needed:
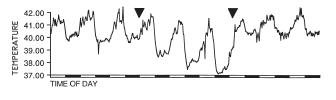
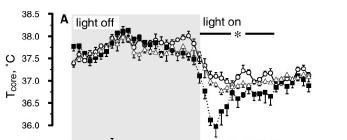
As stated above, hypothermia occurs only during the night while the temperature during the days remains normal. However, all the studies are performed either in natural conditions of day and night or in light-dark cycles in the laboratory. In constant darkness, the circadian rhythm of temperature persists and hypothermia is apparent. Moreover, the temperature drops both at the minima during the 'subjective night' and at the maxima during the 'subjective day' (from [17]):
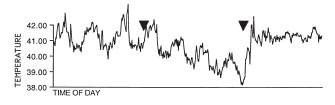
This suggests that light has a direct (or "
masking") effect on body temperature during the light-phase of the cycle. But is this effect acting directly on the thermoregulatory centers in the hypothalamus or is it mediated by the circadian clock that drives the rhythm of body temperature? In
Japanese quail, the circadian pacemakers are located in the eyes. When the eyes are removed [17], both the daily maxima in the light-phase and the nightly minima during the dark phase drop, suggesting that the effect is mediated via the circadian clock, as the light perceived by the photoreceptors in the pineal gland and in the deep brain is incapable of keeping the daily maxima from dropping:
 Mammals
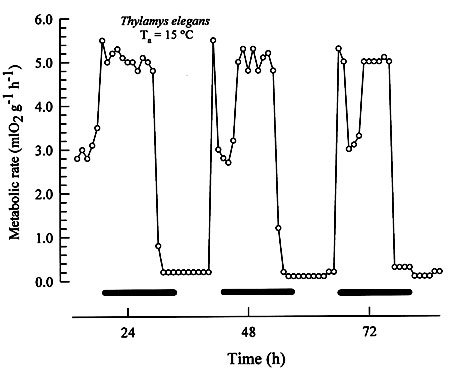
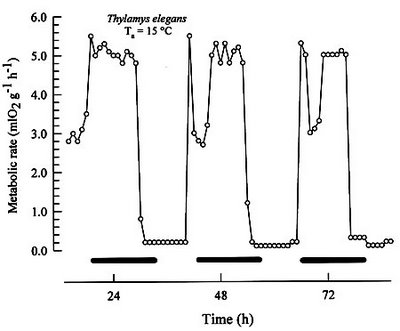
MammalsSome small mammals, such as smallest rodents and shrews, exhibit a full-blown daily torpor either normally [21] or in response to fasting [22]. Here is an example of a daily torpor of a mouse-opposum:
In nocturnal animals, which many mammals are, body temperature is high at night when the animals are active and it drops during the day when the animals are sleeping. In rats, fasting induces diurnal hypothermia, i.e., drop of the daily minimum during the day (black circles, compared to pre- and post- treatment values in white symbols) while the nightly maxima remain unaffected [23]:
Chronic caloric food restriction leads to the drop in both the daily minima and nightly maxima of temperature [24].

All the studies until recently have studied responses in relatively small animals (both birds and mammals) with high metabolic rates and high energy needs. But do larger animals, like humans, also exhibit Chossat's effect? After all, the first documented case, that by Chossat himself, was in a dog. This was repeated recently [25]. But even dogs are pretty small compared to humans.
Recently, researchers have addressed this question in a number of species of large mammals, including sheep, goats, horses and yaks [26-29]. Some additional environmental cues were also studied, including the effects of shearing on the circadian temperature rhythm in sheep [30]. Here is a record from a goat:

Notice that, unlike in birds, both the maxima and minima gradually go down.
But, as far as I could find by digging through the literature, nobody has ever performed a similar study in humans. I am assuming that it has been noticed if body temperature drops in fasted humans, but I am not aware of a study systematically addressing this question.
HumansA few years ago I was teaching one of many sections of an Animal Anatomy and Physiology Course. This course requires students to perform a research project. One group of students studied the effects of fasting on body temperature and blood pressure in humans.
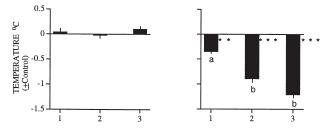
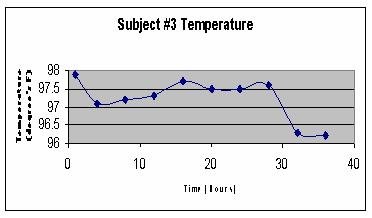
They found 8 subjects, all healthy, athletic, non-drinking, non-smoking students ages 19-23. They were instructed to eat normally during the Day1 of the experiment. They subequently spent 36 hours in a house drinking only water and eating nothing. Every four hours, temperature and pressure were measured. By using kids' digital ear thermometers and manual sphigmomanometers they managed, for the most part, not to awaken the subjects during the night. Here are examples of body temperature of three of the subjects - Night1, followed by Day2 and Night 2:
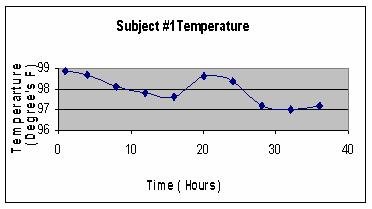

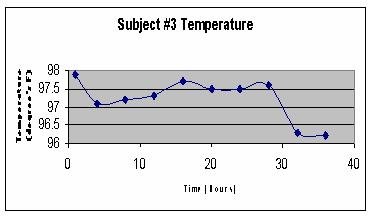
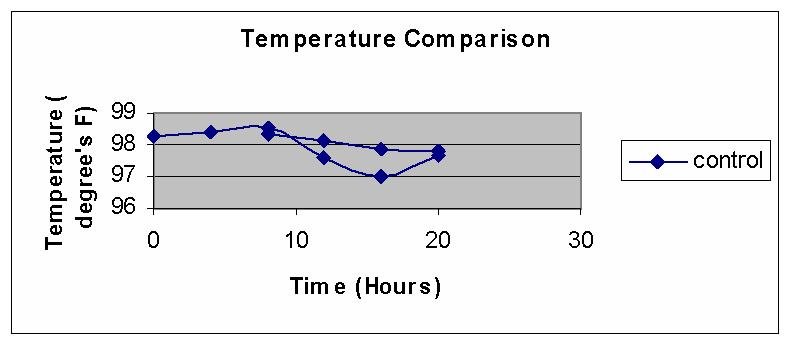
Here are the pooled data for all eight subjects, starting with Day2 and followed by Night1 and Night2 plotted on top of each other for comparison:

Obviously, body temperature of Night2, after a day of fasting, was lower than that of Night1, after the day of normal feeding. I do not have their raw data any more, but if I remember correctly, the data for blood pressure looked very similar. I heard they had a huge breakfast, courtesy of the young researchers, at the end of the experiment.
So, Chossat's Effect appears to be operating in humans as well. Now, this is cool in itself, and I sure hope that someone with access to good clinical lab repeats this study, but there is something else about these data that really excites me. This finding can be used as a tool for studying something entirely different!
The HypothesisOne of the first demonstrations that humans have daily rhythms involved the time-of-day dependence of time perception. In other words, our subjective "feel" of the speed of passage of time changes systematically with the time of day. At the same time, it has been known for a couple of centuries now that the subjective time perception is also altered during fever. And we know that circadian clock governs daily rhtyhms of body temperature. So, what affects the time perception: time of day or body temperature? If the time passes faster in the evening than at dawn, is it because of the circadian clock acting on the time-perception brain-centers directly, or because we are warmer at the time (which is also driven by the circadian clock)?
This question has haunted circadian researchers for decades and they have devised ever more elaborate experiments to tease the two hypotheses apart, with no avail - we still do not know. But, if by depriving the subjects of food, we can dissociate clock-time from temperature, perhaps we can address this question after all. If the subjective perception of 1 minute (do not use 1 second or 1 hour - those are durations unsuited for this experiment) is similar between the night after a fed day and the night after the fasting day, then the perception is directly driven by the circadian clock. If, on the other hand, perception of a minute changes systematically between the two nights, then we conclude that it is body temperature that affects subjective time perception. Please, someone do this! And if you do, or even if you just want to replicate the Chossat's Effect in humans, I would appreciate it if you would
properly cite this post:
Bora Zivkovic, Chossat's Effect in humans and other animals (April 9, 2006), blog Circadiana, http://circadiana.blogspot.com/2006/04/chossats-effect-in-humans-and-other.html
REFERENCES:
[1] M. Chossat, Sur l'inanition, Paris, 1843
[2] Hiebert, S.M. 1990. Energy costs and temporal organization of torpor in the rufous hummingbird (Selasphorus rufus). Physiological Zoology . 63:1082-1097.
[3] Hiebert, S.M. 1991. Seasonal differences in the response of rufous hummingbirds to food restriction: body mass and the use of torpor. Condor 93:526-537.
[4] Tobias Wang, Carrie C.Y. Hung, David J. Randall, THE COMPARATIVE PHYSIOLOGY OF FOOD DEPRIVATION: From Feast to Famine, Annual Review of Physiology, January 2006, Vol. 68, Pages 223-251
[5] Wingfield, JC; Maney, DL; Breuner, CW; Jacobs, JD; Lynn, S; Ramenofsky, M; Richardson, RD, Ecological bases of hormone-behavior interactions: The "emergency life history stage", American Zoologist [Am. Zool.]. Vol. 38, no. 1, pp. 191-206. 1998.
[6] Tracy A. Maddocks, Fritz Geiser, Energetics, Thermoregulation and Nocturnal Hypothermia in Australian Silvereyes, Condor, Vol. 99, No. 1 (Feb., 1997) , pp. 104-112
[7] Randi Eidsmo Reinertsen and Svein Haftorn, The effect of short-time fasting on metabolism and nocturnal hypothermia in the Willow Tit Parus montanus, Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, Volume 154, Number 1 (January 1984): 23 - 28
[8] Barry G. Lovegrove and Gary A. Smith, Is 'nocturnal hypothermia' a valid physiological concept in small birds?: a study on Bronze Mannikins Spermestes cucullatus, Ibis, Volume 145, Issue 4, Page 547 - October 2003
[9] MacMillen RE, Trost CH., Nocturnal hypothermia in the Inca dove, Scardafella inca, Comp Biochem Physiol. 1967 Oct;23(1):243-53.
[10] Colleen T. Downs, Mark Brown, NOCTURNAL HETEROTHERMY AND TORPOR IN THE MALACHITE SUNBIRD (NECTARINIA FAMOSA).
[11] Waite, TA, Nocturnal hypothermia in gray jays Perisoreus canadensis wintering in interior Alaska, ORNIS SCAND. Vol. 22, no. 2, pp. 107-110. 1991.
[12] Cécile Thouzeau, Claude Duchamp, and Yves Handrich, Energy Metabolism and Body Temperature of Barn Owls Fasting in the Cold, Physiological and Biochemical Zoology, volume 72 (1999), pages 170–178
[13] Rashotte ME, Henderson D., Coping with rising food costs in a closed economy: feeding behavior and nocturnal hypothermia in pigeons, J Exp Anal Behav. 1988 Nov;50(3):441-56.
[14] R. Graf, S. Krishna and H. C. Heller, Regulated nocturnal hypothermia induced in pigeons by food deprivation, Am J Physiol Regul Integr Comp Physiol 256: R733-R738, 1989
[15] Michael E. Rashotte, Iuri F. Pastukhov, Eugene L. Poliakov, and Ross P. Henderson, Vigilance states and body temperature during the circadian cycle in fed and fasted pigeons (Columba livia), Am J Physiol Regul Integr Comp Physiol 275: R1690-R1702, 1998
[16] Hohtola E, Hissa R, Pyornila A, Rintamaki H, Saarela S., Nocturnal hypothermia in fasting Japanese quail: the effect of ambient temperature, Physiol Behav. 1991 Mar;49(3):563-7.
[17] Herbert Underwood, Christopher T. Steele and Bora Zivkovic, Effects of Fasting on the Circadian Body Temperature Rhythm of Japanese Quail, Physiology & Behavior, Vol. 66, No. 1, pp. 137–143, 1999
[18] Mirja Laurila, Tiina Pilto, Esa Hohtola, Testing the flexibility of fasting-induced hypometabolism in birds: effect of photoperiod and repeated food deprivations, Journal of Thermal Biology 30 (2005) 131–138
[19] MIRJA LAURILA, THERMOREGULATORY CONSEQUENCES OF STARVATION AND DIGESTION IN BIRDS, PhD Dissertation, Faculty of Science, Department of Biology, University of Oulu, 2005 (http://herkules.oulu.fi/isbn9514277147/isbn9514277147.pdf)
[20] Mirja Laurila, Esa Hohtola, The effect of ambient temperature and simulated predation risk on fasting-induced nocturnal hypothermia of pigeons in outdoor conditions, Journal of Thermal Biology 30 (2005) 392–399
[21] Francisco Bozinovic, Gricelda RuÍz, Arturo CortÉs & Mario Rosenmann, Energetics, thermoregulation and torpor in the Chilean mouse-opossum Thylamys elegans (Didelphidae), Revista Chilena de Historia Natural 78: 199-206, 2005
[22] Lovegrove BG, Raman J, Perrin MR., Daily torpor in elephant shrews (Macroscelidea: Elephantulus spp.) in response to food deprivation, J Comp Physiol [B]. 2001 Feb;171(1):11-21.
[23] Kei Nagashima, Sadamu Nakai, Kenta Matsue, Masahiro Konishi, Mutsumi Tanaka, and Kazuyuki Kanosue, Effects of fasting on thermoregulatory processes and the daily oscillations in rats, Am J Physiol Regul Integr Comp Physiol 284: R1486–R1493, 2003.
[24] Yoda T, Crawshaw LI, Yoshida K, Su L, Hosono T, Shido O, Sakudara S, Fukuda Y & Kanosue K (2000) Effects of food deprivation on daily changes in body temperature and behavioural thermoregulation in rats. Am J Physiol 278: R134-R139.
[25] G. Piccione, G. Caola and R. Refinetti, Daily Rhythms of Blood Pressure, Heart Rate, and Body Temperature in Fed and Fasted Male Dogs, J. Vet. Med. A 52, 377–381 (2005)
[26] Giuseppe Piccione, Giovanni Caola, Roberto Refinetti, Circadian rhythms of body temperature and liver function in fed and food-deprived goats, Comparative Biochemistry and Physiology Part A 134 (2003) 563–572
[27] Piccione, G., Caola, G., Refinetti, R., 2002a. Circadian modulation of starvation-induced hypothermia in sheep and goats. Chronobiol. Int. 19, 531–541.
[28] Piccione, G., Caola, G., Refinetti, R., 2002b. The circadian rhythm of body temperature of the horse. Biol. Rhythm Res. 33, 113–119.
[29] Xing-Tai Han, Ao-Yun Xie, Xi-Chao Bi, Shu-Jie Liu and Ling-Hao Hu, Effects of high altitude and season on fasting heat production in the yak Bos grunniens or Poephagus grunniens, British Journal of Nutrition (2002), 88, 189–197
[30] Giuseppe Piccione, Giovanni Caola, and Roberto Refinetti, Effect of shearing on the core body temperature of three breeds of Mediterranean sheep, Small Ruminant Research 46 (2002) 211–215